뉴런
뉴런(미국 영어: neuron), 뉴론(영국 영어: neurone),[^32] 또는 신경 세포는 흥분성을 가진 세포로, 신경계, 주로 중추신경계의 신경망을 통해 활동 전위라 불리는 전기 신호를 발생시키며, 자극을 수신하고 전달하는 데 도움을 준다. 뉴런은 시냅스를 통해 다른 세포와 소통하는데, 시냅스는 일반적으로 미량의 화학적 신경전달물질을 사용하여 시냅스 전 뉴런에서 시냅스 틈을 거쳐 표적 세포로 전기 신호를 전달하는 특수한 연결 구조이다.
뉴런은 해면동물과 판형동물을 제외한 모든 동물의 신경 조직을 구성하는 주요 요소이다. 식물과 균류에는 신경 세포가 없다. 분자 수준의 증거에 따르면 전기 신호를 생성하는 능력은 약 7억~8억 년 전 토니아기에 진화 과정에서 처음 나타났다. 뉴런의 전신은 펩티드성 분비 세포였다. 이 세포들은 결국 시냅스 후 스캐폴드와 빠른 전기 신호를 생성하는 이온 채널을 만들 수 있게 해 주는 새로운 유전자 모듈을 획득하였다. 전기 신호를 생성하는 능력은 신경계 진화에서 핵심적인 혁신이었다.[^33]
뉴런은 일반적으로 기능에 따라 세 가지 유형으로 분류된다. 감각 뉴런은 감각 기관의 세포에 영향을 미치는 촉각, 소리, 빛 등의 자극에 반응하며, 척수를 거쳐 뇌의 감각 영역으로 신호를 보낸다. 운동 뉴런은 뇌와 척수로부터 신호를 받아 근육 수축[^34]에서 분비샘 활동에 이르기까지 모든 것을 제어한다. 개재 뉴런은 뇌 또는 척수의 같은 영역 내에서 뉴런을 다른 뉴런과 연결한다. 여러 뉴런이 기능적으로 함께 연결되면 신경 회로라 불리는 것을 형성한다.
뉴런은 핵, 미토콘드리아, 골지체 등 다른 세포와 동일한 구조를 모두 포함하고 있지만, 축삭과 수상돌기 같은 고유한 추가 구조를 가지고 있다.[^5] 세포체(소마)는 밀집된 구조이며, 축삭과 수상돌기는 세포체에서 뻗어 나오는 돌기이다. 수상돌기는 일반적으로 풍부하게 분지하며 세포체에서 수백 마이크로미터까지 뻗어 있다. 축삭은 축삭 소구라 불리는 팽대부에서 세포체를 떠나 인간의 경우 최대 1미터, 다른 종에서는 그 이상까지 뻗어 나간다. 축삭은 분지하지만 보통 일정한 직경을 유지한다. 축삭 가지의 가장 먼 끝에는 축삭 말단이 있으며, 여기서 뉴런은 시냅스를 통해 다른 세포로 신호를 전달할 수 있다. 뉴런은 수상돌기가 없거나 축삭이 없을 수도 있다. 신경돌기(neurite)라는 용어는 수상돌기 또는 축삭을 지칭하는 데 사용되며, 특히 세포가 미분화 상태일 때 사용된다.
대부분의 뉴런은 수상돌기와 세포체를 통해 신호를 수신하고 축삭을 따라 신호를 내보낸다. 대다수의 시냅스에서 신호는 한 뉴런의 축삭에서 다른 뉴런의 수상돌기로 전달된다. 그러나 시냅스는 축삭과 다른 축삭, 또는 수상돌기와 다른 수상돌기를 연결할 수도 있다. 신호 전달 과정은 부분적으로 전기적이고 부분적으로 화학적이다. 뉴런은 세포막 양쪽의 전압 기울기가 유지되기 때문에 전기적으로 흥분성을 가진다. 짧은 시간 내에 전압이 충분히 크게 변하면 뉴런은 활동 전위라 불리는 전부 아니면 전무(all-or-nothing) 방식의 전기화학적 펄스를 생성한다. 이 전위는 축삭을 따라 빠르게 이동하며 도달하는 곳의 시냅스 연결을 활성화한다. 시냅스 신호는 흥분성 또는 억제성일 수 있으며, 세포체에 도달하는 순 전압을 증가시키거나 감소시킨다.
대부분의 경우 뉴런은 뇌 발달 과정과 유아기에 신경 줄기세포에 의해 생성된다. 신경 발생은 뇌의 대부분 영역에서 성인기에 접어들면 대체로 중단된다.
신경계
신경세포는 구조적·대사적 지지를 제공하는 신경아교세포와 함께 신경계의 주요 구성 요소이다.[^1] 신경계는 뇌와 척수를 포함하는 중추신경계와 자율신경계, 장신경계, 체성신경계를 포함하는 말초신경계로 구성된다.[^35] 척추동물에서 대부분의 신경세포는 중추신경계에 속하지만, 일부는 말초 신경절에 존재하며, 많은 감각 신경세포는 망막이나 달팽이관과 같은 감각 기관에 위치한다.
축삭은 다발을 이루어 말초신경계의 신경을 구성할 수 있다(마치 여러 가닥의 전선이 모여 케이블을 이루는 것과 같다). 중추신경계에서 축삭 다발은 신경로라고 불린다.
해부학 및 조직학
뉴런은 세포 신호의 처리와 전달에 고도로 특화되어 있다. 신경계의 각 부위에서 수행하는 기능이 다양한 만큼, 뉴런의 형태, 크기, 전기화학적 특성도 매우 다양하다. 예를 들어, 뉴런의 세포체 직경은 4~100마이크로미터까지 다양할 수 있다.[^36]
- 세포체(soma)는 뉴런의 몸체이다. 핵을 포함하고 있기 때문에 대부분의 단백질 합성이 이곳에서 일어난다. 핵의 직경은 3~18마이크로미터 범위이다.[^37]
- 뉴런의 수상돌기(dendrite)는 많은 가지를 가진 세포 돌기이다. 이러한 전체적인 형태와 구조를 비유적으로 수상돌기 나무(dendritic tree)라고 한다. 가지들은 여러 크기 규모에서 반복되는 프랙탈 패턴을 형성한다.[^38] 이 프랙탈 나무 구조는 수상돌기 가시(dendritic spine)를 통해 뉴런에 대한 입력의 대부분이 이루어지는 곳이다.
- 축삭(axon)은 더 가늘고 케이블과 같은 돌기로, 세포체 직경의 수십, 수백, 심지어 수만 배 길이로 뻗어나갈 수 있다. 축삭은 주로 세포체에서 멀어지는 방향으로 신경 신호를 전달하며, 일부 유형의 정보를 세포체로 다시 전달하기도 한다. 많은 뉴런은 하나의 축삭만 가지고 있지만, 이 축삭은 광범위하게 분지하여 많은 표적 세포와의 소통을 가능하게 한다. 축삭이 세포체에서 나오는 부분을 축삭 언덕(axon hillock)이라고 한다. 해부학적 구조일 뿐만 아니라, 축삭 언덕은 전압 의존성 나트륨 채널의 밀도가 가장 높은 곳이기도 하다. 이로 인해 뉴런에서 가장 쉽게 흥분하는 부분이자 축삭의 스파이크 발생 구역이 된다. 전기생리학적으로 가장 음의 역치 전위를 가진다. ** 축삭과 축삭 언덕은 일반적으로 정보 유출에 관여하지만, 이 영역은 다른 뉴런으로부터 입력을 받을 수도 있다.
- 축삭 말단(axon terminal)은 세포체에서 가장 먼 축삭 끝에 위치하며 시냅스를 포함한다. 시냅스 종말 단추(synaptic bouton)는 표적 뉴런과 소통하기 위해 신경전달물질 화학물질이 방출되는 특화된 구조이다. 축삭 말단의 시냅스 종말 단추 외에도, 뉴런은 축삭 길이를 따라 위치하는 통과 종말 단추(en passant bouton)를 가질 수 있다.
일반적으로 받아들여지는 뉴런에 대한 견해는 각 해부학적 구성요소에 고유한 기능을 부여하지만, 수상돌기와 축삭은 소위 주요 기능과 반대되는 방식으로 작용하는 경우가 많다.
중추신경계의 축삭과 수상돌기는 일반적으로 약 1마이크로미터 두께에 불과하지만, 말초신경계의 일부는 훨씬 더 두껍다. 세포체는 보통 직경이 약 10~25마이크로미터이며, 그 안에 포함된 세포핵보다 그다지 크지 않은 경우가 많다. 인간 운동 뉴런의 가장 긴 축삭은 1미터 이상으로, 척추 하부에서 발가락까지 도달할 수 있다.
감각 뉴런은 발가락에서 척수 후주까지 이어지는 축삭을 가질 수 있으며, 성인에서 1.5미터 이상이다. 기린은 목 전체 길이를 따라 수 미터 길이의 단일 축삭을 가지고 있다. 축삭 기능에 대해 알려진 많은 것은 오징어 거대 축삭을 연구하여 얻은 것으로, 비교적 거대한 크기(두께 0.5~1밀리미터, 길이 수 센티미터) 덕분에 이상적인 실험 재료이다.
완전히 분화된 뉴런은 영구적으로 유사분열 후기 상태이지만,[^39] 성인 뇌에 존재하는 줄기세포가 유기체의 일생에 걸쳐 기능적 뉴런을 재생할 수 있다(신경발생 참조). 별아교세포(astrocyte)는 별 모양의 아교세포로, 만능성이라는 줄기세포와 유사한 특성에 의해 뉴런으로 전환되는 것이 관찰되었다.[^2]
세포막
모든 동물 세포와 마찬가지로, 모든 뉴런의 세포체는 많은 종류의 내장 단백질 구조를 가진 지질 이중층인 원형질막으로 둘러싸여 있다.[^40] 지질 이중층은 강력한 전기 절연체이지만, 뉴런에서는 막에 내장된 많은 단백질 구조가 전기적으로 활성화되어 있다. 여기에는 전하를 띤 이온이 막을 가로질러 흐르게 하는 이온 채널과 막의 한쪽에서 다른 쪽으로 이온을 화학적으로 수송하는 이온 펌프가 포함된다. 대부분의 이온 채널은 게이트가 있어 특정 유형의 이온에만 투과성을 보인다. 일부 이온 채널은 전압 개폐성으로, 막을 가로지르는 전압 차이를 변경하여 열림과 닫힘 상태를 전환할 수 있다. 다른 것들은 화학적 개폐성으로, 세포외액을 통해 확산되는 화학물질과의 상호작용에 의해 열림과 닫힘 상태를 전환할 수 있다. 관련 이온에는 나트륨, 칼륨, 염소, 칼슘이 포함된다. 이온 채널과 이온 펌프 사이의 상호작용은 막을 가로지르는 전압 차이를 생성하며, 기저 상태에서 일반적으로 1/10볼트보다 약간 작다. 이 전압은 두 가지 기능을 가진다: 첫째, 막에 내장된 다양한 전압 의존성 단백질 기구에 전원을 공급하고; 둘째, 막의 서로 다른 부분 사이의 전기 신호 전달을 위한 기반을 제공한다.
조직학 및 내부 구조
 인간 해마 조직의 뉴런]]
인간 해마 조직의 뉴런]]

신경 세포체를 호염기성("염기를 좋아하는") 염료로 염색하면 니슬 소체(또는 니슬 물질)라고 불리는 수많은 미세한 덩어리가 관찰된다. 이 구조는 조면소포체와 관련 리보솜 RNA로 구성되어 있다. 독일의 정신과 의사이자 신경병리학자 Franz Nissl(1860~1919)의 이름을 딴 이 소체는 단백질 합성에 관여하며, 신경세포가 대사적으로 매우 활발하다는 사실로 그 풍부함을 설명할 수 있다. 아닐린이나 (약하게) 헤마톡실린[^41] 같은 호염기성 염료는 음전하를 띤 성분을 강조하여, 리보솜 RNA의 인산 골격에 결합한다.
뉴런의 세포체는 신경미세섬유(neurofilament)라 불리는 복잡한 구조 단백질 그물에 의해 지지되며, 이는 신경미세관(neurotubule, 뉴런 미세소관)과 함께 더 큰 신경원섬유(neurofibril)로 조립된다.[^3] 일부 뉴런은 신경멜라닌(카테콜아민 합성의 부산물인 갈흑색 색소)과 리포푸신(황갈색 색소) 같은 색소 과립도 포함하며, 이 두 가지 모두 나이가 들면서 축적된다.[^42][^43][^44] 뉴런 기능에 중요한 다른 구조 단백질로는 액틴과 미세소관의 튜불린이 있다. 제III형 β-튜불린은 거의 뉴런에서만 발견된다. 액틴은 주로 뉴런 발달 중에 축삭과 수상돌기의 끝부분에서 발견된다. 그곳에서 액틴 역학은 미세소관과의 상호작용을 통해 조절될 수 있다.[^45]
축삭과 수상돌기 사이에는 서로 다른 내부 구조적 특성이 있다. 전형적인 축삭은 초기 분절 일부를 제외하면 리보솜을 거의 포함하지 않는다. 수상돌기는 과립형 소포체 또는 리보솜을 포함하며, 세포체에서 멀어질수록 그 양이 감소한다.
분류
![Image of pyramidal neurons in mouse [cerebral cortex expressing green fluorescent protein. The red staining indicates GABAergic interneurons.[^46]]] 신경세포는 형태와 크기가 다양하며, 형태학적 특성과 기능에 따라 분류할 수 있다.[^4] 해부학자 카밀로 골지(Camillo Golgi)는 신경세포를 두 가지 유형으로 분류하였다. 제1형은 장거리 신호 전달에 사용되는 긴 축삭돌기를 가진 것이고, 제2형은 짧은 축삭돌기를 가진 것으로, 종종 수상돌기와 혼동될 수 있다. 제1형 세포는 세포체의 위치에 따라 더 세분화할 수 있다. 척수 운동 신경세포로 대표되는 제1형 신경세포의 기본 형태는 세포체(소마)와 수초로 둘러싸인 길고 가는 축삭돌기로 구성된다. 수상돌기 나무는 세포체를 감싸며 다른 신경세포로부터 신호를 수신한다. 축삭돌기의 끝에는 분지된 축삭 말단이 있으며, 이 말단과 다음 신경세포의 수상돌기 사이에 있는 시냅스 틈이라는 간격으로 신경전달물질을 방출한다.
{kind=link}
구조적 분류
극성

대부분의 신경세포는 해부학적으로 다음과 같이 특성화할 수 있다:[^5]
- 단극 신경세포: 하나의 돌기를 가진다. 단극 세포는 전적으로 감각 신경세포이다. 수상돌기가 감각 정보를 수신하며, 때로는 자극 자체로부터 직접 수신하기도 한다. 단극 신경세포의 세포체는 항상 신경절에 위치한다. 감각 수용은 말초 기능이므로 세포체는 말초에 위치하지만, 신경절 내에서 중추신경계에 더 가까이 있다. 축삭돌기는 수상돌기 말단에서 시작하여 신경절의 세포체를 지나 중추신경계로 투사된다.
- 양극 신경세포: 축삭돌기 1개와 수상돌기 1개를 가진다. 주로 후각 상피와 망막의 일부에서 발견된다.
- 다극 신경세포: 축삭돌기 1개와 수상돌기 2개 이상을 가진다 ** 골지 I형: 장거리 투사 축삭돌기를 가진 신경세포; 예로는 피라미드 세포, 푸르킨예 세포, 전각 세포가 있다 ** 골지 II형: 축삭돌기가 국소적으로 투사되는 신경세포; 가장 좋은 예는 과립 세포이다
- 무축삭 신경세포: 축삭돌기를 수상돌기와 구별할 수 없는 경우
- 위단극 신경세포: 하나의 돌기가 축삭돌기와 수상돌기 역할을 동시에 수행한다
기타
일부 고유한 신경세포 유형은 신경계 내 위치와 독특한 형태에 따라 식별할 수 있다. 몇 가지 예는 다음과 같다:
- 바구니 세포, 피질과 소뇌에서 발견되며 표적 세포의 세포체 주위에 조밀한 말단 신경총을 형성하는 개재 신경세포
- 베츠 세포, 일차 운동 피질의 큰 운동 신경세포
- 루가로 세포, 소뇌의 개재 신경세포
- 중형 가시 신경세포, 선조체에서 대부분을 차지하는 신경세포
- 푸르킨예 세포, 소뇌의 거대한 신경세포로, 골지 I형 다극 신경세포의 한 유형
- 피라미드 세포, 삼각형 세포체를 가진 신경세포로, 골지 I형의 한 유형
- 로즈힙 세포, 피라미드 세포와 상호 연결되는 인간 고유의 억제성 신경세포
- 렌쇼 세포, 양쪽 끝이 알파 운동 신경세포에 연결된 신경세포
- 단극 브러시 세포, 브러시 모양의 술 형태로 끝나는 독특한 수상돌기를 가진 개재 신경세포
- 과립 세포, 골지 II형 신경세포의 한 유형
- 전각 세포, 척수에 위치한 운동 신경세포
- 방추 세포, 뇌의 널리 분리된 영역을 연결하는 개재 신경세포
기능적 분류
방향
- 구심성 신경세포는 조직과 기관에서 중추신경계로 정보를 전달하며, 감각 신경세포라고도 불린다.
- 원심성 신경세포(운동 신경세포)는 중추신경계에서 효과기 세포로 신호를 전달한다.
- 개재 신경세포는 중추신경계의 특정 영역 내에서 신경세포를 연결한다.
구심성과 원심성은 일반적으로 각각 뇌로 정보를 가져오거나 뇌에서 정보를 보내는 신경세포를 가리키기도 한다.
다른 신경세포에 대한 작용
신경세포는 화학적 수용체에 결합하는 신경전달물질을 방출하여 다른 신경세포에 영향을 미친다. 시냅스 후 신경세포에 대한 효과는 시냅스 전 신경세포나 신경전달물질이 아닌, 활성화되는 수용체의 유형에 의해 결정된다. 수용체는 크게 흥분성 (발화율 증가 유발), 억제성 (발화율 감소 유발), 또는 조절성 (발화율과 직접 관련되지 않는 장기 지속 효과 유발)으로 분류된다.
뇌에서 가장 흔한(90% 이상) 두 가지 신경전달물질인 글루타메이트와 GABA는 대체로 일관된 작용을 한다. 글루타메이트는 여러 유형의 수용체에 작용하며, 이온성 수용체에서는 흥분성 효과를, 대사성 수용체에서는 조절성 효과를 나타낸다. 마찬가지로 GABA도 여러 유형의 수용체에 작용하지만, 모든 수용체에서 억제성 효과를 나타낸다(적어도 성체 동물에서는). 이러한 일관성 때문에 신경과학자들은 글루타메이트를 방출하는 세포를 "흥분성 신경세포", GABA를 방출하는 세포를 "억제성 신경세포"라고 부르는 것이 일반적이다. 일부 다른 유형의 신경세포도 일관된 효과를 가지는데, 예를 들어 척수에서 아세틸콜린을 방출하는 "흥분성" 운동 신경세포와 글리신을 방출하는 "억제성" 척수 신경세포가 있다.
흥분성 신경전달물질과 억제성 신경전달물질의 구분은 절대적이지 않다. 오히려 이는 시냅스 후 신경세포에 존재하는 화학적 수용체의 종류에 따라 달라진다. 원칙적으로, 단일 신경전달물질을 방출하는 단일 신경세포가 일부 표적에는 흥분성 효과를, 다른 표적에는 억제성 효과를, 또 다른 표적에는 조절성 효과를 미칠 수 있다. 예를 들어, 망막의 광수용체 세포는 빛이 없을 때 지속적으로 신경전달물질 글루타메이트를 방출한다. 소위 OFF 양극 세포는 대부분의 신경세포와 마찬가지로 방출된 글루타메이트에 의해 흥분된다. 그러나 인접한 표적 신경세포인 ON 양극 세포는 전형적인 이온성 글루타메이트 수용체가 없고 대신 억제성 대사성 글루타메이트 수용체를 발현하기 때문에, 오히려 글루타메이트에 의해 억제된다.[^47] 빛이 존재하면 광수용체는 글루타메이트 방출을 중단하고, 이로 인해 ON 양극 세포가 억제에서 해방되어 활성화된다. 동시에 이는 OFF 양극 세포에서 흥분을 제거하여 침묵시킨다.
시냅스 전 신경세포가 발현하는 단백질을 기반으로 시냅스 후 신경세포에 미칠 억제 효과의 유형을 식별하는 것이 가능하다. 파르브알부민 발현 신경세포는 일반적으로 시각 피질에서 시냅스 후 신경세포의 출력 신호를 감쇠시키는 반면, 소마토스타틴 발현 신경세포는 일반적으로 시냅스 후 신경세포에 대한 수상돌기 입력을 차단한다.[^6]
발화 패턴
신경세포는 내재적 막전위 진동 패턴과 같은 고유한 전기반응 특성을 가진다.[^7] 따라서 신경세포는 전기생리학적 특성에 따라 분류할 수 있다:
- 지속성 또는 규칙적 스파이킹. 일부 신경세포는 전형적으로 일정한 주파수로 지속적(긴장성)으로 활동한다. 예: 신선조체의 개재 신경세포.
- 위상성 또는 버스트형. 버스트로 발화하는 신경세포를 위상성이라 한다.
- 빠른 스파이킹. 일부 신경세포는 높은 발화율로 주목받는데, 예를 들어 일부 유형의 피질 억제성 개재 신경세포, 담창구 세포, 망막 신경절 세포가 있다.[^48][^49]
신경전달물질

신경전달물질은 하나의 신경세포에서 다른 신경세포, 근육 세포 또는 분비샘 세포로 전달되는 화학적 전달물질이다.
-
콜린성 신경세포 – 아세틸콜린. 아세틸콜린은 시냅스 전 신경세포에서 시냅스 틈으로 방출된다. 이는 리간드 개폐 이온 통로와 대사성(GPCR) 무스카린 수용체 모두의 리간드로 작용한다. 니코틴 수용체는 니코틴에 결합하는 알파 및 베타 소단위로 구성된 오량체 리간드 개폐 이온 통로이다. 리간드 결합은 통로를 열어 Na+ 유입에 의한 탈분극을 일으키고 시냅스 전 신경전달물질 방출 확률을 증가시킨다. 아세틸콜린은 콜린과 아세틸 조효소 A로부터 합성된다.
-
아드레날린성 신경세포 – 노르아드레날린. 노르아드레날린(노르에피네프린)은 교감신경계의 대부분의 절후 신경세포에서 두 가지 GPCR 세트, 즉 알파 아드레날린 수용체와 베타 아드레날린 수용체로 방출된다. 노르아드레날린은 세 가지 일반적인 카테콜아민 신경전달물질 중 하나이며, 말초신경계에서 가장 흔하다. 다른 카테콜아민과 마찬가지로 티로신에서 합성된다.
-
GABA성 신경세포 – 감마 아미노부티르산. GABA는 글리신과 함께 중추신경계(CNS)의 두 가지 신경 억제 물질 중 하나이다. GABA는 ACh와 유사한 기능을 하여, Cl− 이온이 시냅스 후 신경세포로 유입될 수 있도록 음이온 통로를 개폐한다. Cl−는 신경세포 내 과분극을 일으켜, 전압이 더 음으로 되면서 활동전위 발화 확률을 감소시킨다(활동전위가 발화하려면 양의 전압 역치에 도달해야 한다). GABA는 글루타메이트 탈탄산효소에 의해 글루타메이트 신경전달물질로부터 합성된다.
-
글루타메이트성 신경세포 – 글루타메이트. 글루타메이트는 아스파르트산과 함께 두 가지 주요 흥분성 아미노산 신경전달물질 중 하나이다. 글루타메이트 수용체는 네 가지 범주로 나뉘는데, 그 중 세 가지는 리간드 개폐 이온 통로이고 하나는 G-단백질 결합 수용체(흔히 GPCR로 불림)이다.
#AMPA와 카이네이트 수용체는 Na+ 투과성 양이온 통로로 기능하며 빠른 흥분성 시냅스 전달을 매개한다. #NMDA 수용체는 Ca2+에 대한 투과성이 더 높은 또 다른 양이온 통로이다. NMDA 수용체의 기능은 통로 내 보조작용제로서 글리신 수용체 결합에 의존한다. NMDA 수용체는 두 리간드가 모두 존재하지 않으면 기능하지 않는다. #대사성 수용체, 즉 GPCR은 시냅스 전달과 시냅스 후 흥분성을 조절한다. : 글루타메이트는 뇌로의 혈류가 차단되면 흥분독성을 일으켜 뇌 손상을 초래할 수 있다. 혈류가 억제되면 시냅스 전 신경세포에서 글루타메이트가 방출되어, 스트레스 조건 밖의 정상 상태보다 더 큰 NMDA 및 AMPA 수용체 활성화를 일으키고, 이로 인해 Ca2+와 Na+가 시냅스 후 신경세포로 과다 유입되어 세포 손상을 초래한다. 글루타메이트는 글루타메이트 합성효소에 의해 아미노산 글루타민으로부터 합성된다.
-
도파민성 신경세포—도파민. 도파민은 D1형(D1 및 D5) Gs 결합 수용체(cAMP와 PKA를 증가시킴)와 D2형(D2, D3, D4) Gi 결합 수용체(cAMP와 PKA를 감소시킴)에 작용하는 신경전달물질이다. 도파민은 기분 및 행동과 관련되며, 시냅스 전 및 시냅스 후 신경전달을 모두 조절한다. 흑질에서의 도파민 신경세포 손실은 파킨슨병과 연관되어 왔다. 도파민은 아미노산 티로신에서 합성된다. 티로신은 티로신 수산화효소에 의해 레보도파(L-DOPA)로 촉매되고, 레보도파는 방향족 아미노산 탈탄산효소에 의해 도파민으로 전환된다.
-
세로토닌성 신경세포—세로토닌. 세로토닌(5-하이드록시트립타민, 5-HT)은 흥분성 또는 억제성으로 작용할 수 있다. 네 가지 5-HT 수용체 분류 중 3개는 GPCR이고 1개는 리간드 개폐 양이온 통로이다. 세로토닌은 트립토판 수산화효소에 의해 트립토판에서 합성되고, 이후 탈탄산효소에 의해 추가로 합성된다. 시냅스 후 신경세포에서의 5-HT 부족은 우울증과 연관되어 왔다. 시냅스 전 세로토닌 수송체를 차단하는 약물이 치료에 사용되는데, 프로작(Prozac)과 졸로프트(Zoloft)가 그 예이다.
-
퓨린성 신경세포—ATP. ATP는 리간드 개폐 이온 통로(P2X 수용체)와 GPCR(P2Y 수용체) 모두에 작용하는 신경전달물질이다. 그러나 ATP는 공동전달물질로서 가장 잘 알려져 있다. 이러한 퓨린성 신호전달은 아데노신과 같은 다른 퓨린에 의해서도 매개될 수 있으며, 아데노신은 특히 P2Y 수용체에 작용한다.
-
히스타민성 신경세포—히스타민. 히스타민은 단일아민 신경전달물질이자 신경조절물질이다. 히스타민 생성 신경세포는 시상하부의 유두체결절핵에서 발견된다.[^50] 히스타민은 각성 및 수면/각성 행동 조절에 관여한다.
다중모델 분류
2012년 이후 세포 및 전산 신경과학 분야에서 뇌의 모든 신경세포와 종 전반에 적용될 수 있는 보편적 신경세포 분류 체계를 만들려는 움직임이 있어 왔다. 이는 모든 신경세포의 세 가지 본질적 특성, 즉 전기생리학, 형태학, 그리고 개별 세포의 전사체를 고려함으로써 이루어진다. 이 분류는 보편적일 뿐만 아니라 성상세포도 분류할 수 있다는 장점이 있다. 세 가지 특성을 동시에 측정할 수 있는 패치-시퀀싱이라는 방법이 앨런 뇌과학연구소(Allen Institute for Brain Science)에서 광범위하게 사용되고 있다.[^51] 2023년에는 가장 최첨단 분자생물학 접근법을 사용한 국제 연구자 협력을 통해 전사, 후성유전학, 기능 수준에서의 성인 및 발달 중인 인간 뇌의 포괄적 세포 지도가 작성되었다.[^52]
연결성
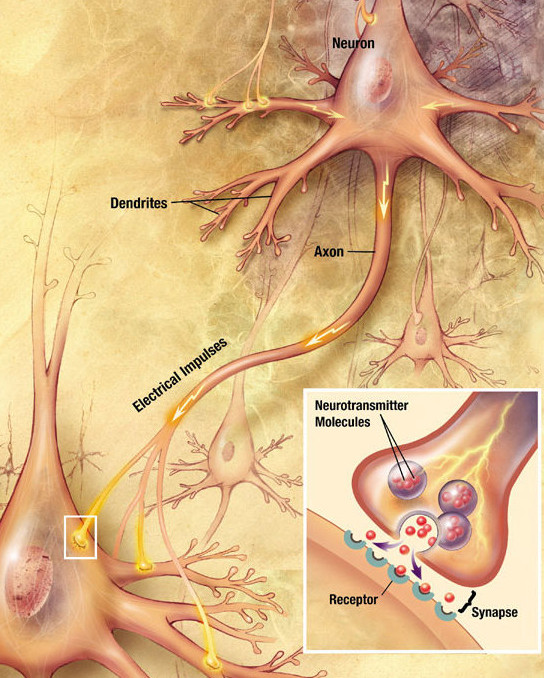

뉴런은 시냅스를 통해 서로 소통하는데, 시냅스에서는 한 세포의 축삭 말단이 다른 뉴런의 수상돌기, 세포체, 또는 드물게는 축삭과 접촉한다. 소뇌의 푸르킨예 세포와 같은 뉴런은 1,000개 이상의 수상돌기 가지를 가지고 있어 수만 개의 다른 세포와 연결을 형성하며, 시상상핵의 대세포 뉴런과 같은 다른 뉴런은 하나 또는 두 개의 수상돌기만을 가지고 있고, 각각의 수상돌기는 수천 개의 시냅스를 받아들인다.
시냅스는 흥분성 또는 억제성일 수 있으며, 각각 표적 뉴런의 활성을 증가시키거나 감소시킨다. 일부 뉴런은 전기적 시냅스를 통해서도 소통하는데, 이는 세포 간의 직접적인 전기 전도성 접합이다.[^53]
활동전위가 축삭 말단에 도달하면 전압 의존성 칼슘 채널이 열리고, 칼슘 이온이 말단으로 유입된다. 칼슘은 신경전달물질 분자로 채워진 시냅스 소포가 세포막과 융합하게 하여 그 내용물을 시냅스 틈으로 방출시킨다. 신경전달물질은 시냅스 틈을 가로질러 확산되어 시냅스후 뉴런의 수용체를 활성화시킨다. 축삭 말단의 높은 세포질 칼슘 농도는 미토콘드리아의 칼슘 흡수를 촉발하며, 이는 다시 미토콘드리아의 에너지 대사를 활성화하여 지속적인 신경전달을 지원하는 ATP를 생산한다.[^8]
자기시냅스는 뉴런의 축삭이 자신의 수상돌기에 연결되는 시냅스이다.
인간의 뇌에는 약 8.6 x 1010개(860억 개)의 뉴런이 있다.[^54][^9] 각 뉴런은 평균적으로 다른 뉴런과 7,000개의 시냅스 연결을 가지고 있다. 3세 아동의 뇌에는 약 1015개(1천조 개)의 시냅스가 있는 것으로 추정된다. 이 수는 나이가 들면서 감소하며, 성인기에 안정화된다. 성인의 경우 추정치는 다양하여 1014개에서 5 x 1014개의 시냅스(100조~500조 개)에 이른다.[^55]
비전기화학적 신호전달
전기적 및 화학적 신호전달 외에도, 연구에 따르면 건강한 인간 뇌의 뉴런은 다음을 통해서도 소통할 수 있다:
- 수상돌기 가시의 확대에 의해 생성되는 힘[^56]
- 단백질의 전달 – 신경세포 간 수송 단백질(TNTPs)[^57][^58]
뉴런은 또한 환경으로부터의 입력과 생물체의 다른 부분에서 분비되는 호르몬에 의해 조절될 수 있으며,[^59] 이는 뉴런에 의해 직간접적으로 영향을 받을 수 있다. 이는 BDNF와 같은 신경영양인자에도 적용된다. 장내 미생물군도 뇌와 연결되어 있다.[^60] 뉴런은 또한 "체세포 접합"이라 불리는 특수한 접촉 부위를 통해 뇌의 주요 면역세포인 미세아교세포와 소통한다. 이러한 연결은 미세아교세포가 뉴런의 기능을 지속적으로 모니터링하고 조절하며, 필요할 때 신경보호 작용을 수행할 수 있게 한다.[^10]
활동전위의 전파 메커니즘
1937년 존 재커리 영은 오징어 거대 축삭이 신경 세포의 전기적 특성을 연구하는 데 사용될 수 있다고 제안하였다.[^61] 이 축삭은 인간의 신경 세포보다 크지만 구조가 유사하여 연구하기가 더 용이하다. 오징어 거대 축삭에 전극을 삽입함으로써 막전위를 정확하게 측정할 수 있었다.
축삭과 세포체의 세포막에는 전압 개폐 이온 통로가 있어 신경 세포가 전기 신호(활동전위)를 생성하고 전파할 수 있게 한다. 일부 신경 세포는 역치 이하의 막전위 진동도 생성한다. 이러한 신호는 나트륨(Na+), 칼륨(K+), 염소(Cl−), 칼슘(Ca2+) 등의 전하 운반 이온에 의해 생성되고 전파된다.
압력, 신장, 화학적 전달 물질, 세포막 전위 변화 등 여러 자극이 신경 세포를 활성화하여 전기적 활동을 유발할 수 있다.[^62] 자극은 세포막 내의 특정 이온 통로를 열리게 하여 세포막을 통한 이온의 흐름을 유발하고, 이로 인해 막전위가 변화한다. 신경 세포는 자신의 신경 세포 유형을 규정하는 특정 전기적 특성을 유지해야 한다.[^11]
가는 신경 세포와 축삭은 활동전위를 생성하고 전달하는 데 대사 비용이 적게 들지만, 굵은 축삭은 신호를 더 빠르게 전달한다. 대사 비용을 최소화하면서 빠른 전도를 유지하기 위해 많은 신경 세포는 축삭 주위에 수초(미엘린)라는 절연 피복을 가지고 있다. 이 피복은 신경아교세포, 즉 중추신경계에서는 희소돌기아교세포가, 말초신경계에서는 슈반세포가 형성한다. 수초는 같은 직경의 무수초 축삭보다 활동전위가 더 빠르게 전달되게 하면서 에너지 소비는 줄여 준다. 말초 신경의 수초는 보통 약 1 mm 길이의 구간으로 축삭을 따라 이어지며, 그 사이사이에 전압 개폐 이온 통로가 고밀도로 분포하는 랑비에 결절이라는 무수초 부분이 있다. 다발성 경화증은 중추신경계 축삭의 탈수초화로 인해 발생하는 신경학적 질환이다.
일부 신경 세포는 활동전위를 생성하지 않고 등급화된 전기 신호를 생성하며, 이것이 등급화된 신경전달물질 방출을 유발한다. 이러한 비발화 신경 세포는 먼 거리까지 신호를 전달할 수 없기 때문에 주로 감각 신경 세포나 사이신경 세포인 경향이 있다.
신경 부호화
신경 부호화는 감각 정보 및 기타 정보가 뇌에서 신경 세포에 의해 어떻게 표상되는지를 다루는 분야이다. 신경 부호화 연구의 주요 목표는 자극과 개별 또는 집단 신경 세포 반응 사이의 관계, 그리고 집단 내 신경 세포들의 전기적 활동 간의 관계를 규명하는 것이다.[^12] 신경 세포는 디지털 정보와 아날로그 정보를 모두 부호화할 수 있는 것으로 여겨진다.[^63]
전무 아니면 전부의 법칙

신경 자극의 전도는 전무 아니면 전부 반응의 한 예이다. 다시 말해, 뉴런이 반응한다면 반드시 완전하게 반응해야 한다. 더 밝은 이미지나 더 큰 소리와 같이 자극의 강도가 더 크더라도 더 강한 신호를 만들어내지는 않지만, 발화 빈도는 증가시킬 수 있다. 수용기는 자극에 대해 다양한 방식으로 반응한다. 느리게 적응하는 수용기 또는 긴장성 수용기는 지속적인 자극에 반응하여 일정한 발화 속도를 유지한다. 긴장성 수용기는 대부분 자극 강도가 증가하면 발화 빈도를 높이는 방식으로 반응하며, 이는 보통 자극 대 초당 자극 횟수를 나타낸 거듭제곱 함수로 표현된다. 이는 특정 주파수(색상)에서 빛의 강도가 높아지려면 더 많은 광자가 필요한 것과 비유할 수 있는데, 특정 주파수에서 광자가 더 "강해질" 수는 없기 때문이다.
다른 수용기 유형에는 빠르게 적응하는 수용기 또는 위상성 수용기가 있으며, 이는 지속적인 자극에 대해 발화가 감소하거나 멈추는 특징이 있다. 예를 들어 피부를 만지면 뉴런이 발화하지만, 물체가 일정한 압력을 유지하면 뉴런은 발화를 멈춘다. 압력과 진동에 반응하는 피부와 근육의 뉴런에는 기능을 돕는 여과 보조 구조가 있다.
파치니 소체는 그러한 구조 중 하나이다. 이것은 양파처럼 동심원 층으로 이루어져 있으며, 축삭 말단 주위에 형성된다. 압력이 가해져 소체가 변형되면 기계적 자극이 축삭에 전달되어 발화가 일어난다. 압력이 일정하게 유지되면 자극은 끝나므로, 이 뉴런들은 보통 초기 변형 시 일시적인 탈분극으로 반응하고, 압력이 제거되어 소체의 형태가 다시 바뀔 때 다시 반응한다. 다른 유형의 적응은 여러 뉴런의 기능을 확장하는 데 중요하다.[^64]
뉴런은 오랫동안 정형화된 최대 반응을 보이거나 전혀 반응하지 않는 것으로 여겨져 왔지만, 이것이 부분적으로만 맞다고 주장하는 연구들이 있다. 뉴런이 활동 전위를 발생시키거나 발생시키지 않는 것은 사실이지만, 뉴런이 발생시키는 활동 전위의 진폭과 지속 시간은 크게 달라질 수 있어, 뉴런이 최소한 활동 전위의 강도로 정보를 부호화할 수 있다는 것이다. 또한 활동 전위에 담긴 아날로그 정보가 원래 불가능하다고 여겨졌던 거리까지 생존하여 전달될 수 있다는 것이 밝혀졌다. 이는 문헌에서 흔히 볼 수 있는 일반적인 발화율 및 시간적 부호화 이론에 비해 매우 효과적인 정보 부호화 방식으로 제안되었으며, 현재의 통념이 시사하는 것보다 약 4배 더 많은 정보를 전달할 수 있는 능력을 가진다.[^65][^66][^67][^68][^69]
어원과 표기
독일의 해부학자 하인리히 빌헬름 발다이어가 1891년에 neuron이라는 용어를 도입했으며,[^17] 이는 고대 그리스어 νεῦρον neuron '힘줄, 끈, 신경'에 기반한다.[^13]
이 단어는 프랑스어에서 neurone이라는 표기로 채택되었다. 영어에서도 많은 저자가 이 표기를 사용했으나,[^14] 현재 미국식 용법에서는 드물게 되었고 영국식 용법에서도 흔하지 않게 되었다.[^15][^13]
일부 이전 문헌에서는 nerve cell(cellule nervose)이라는 용어를 사용했는데, 이는 카밀로 골지가 1873년에 광학 현미경으로 신경 조직을 시각화하는 데 사용되는 은 염색 기법의 발견에 관한 논문에서 채택한 것이다.[^70]
역사
![질산은법으로 염색한 [해마 의 카밀로 골지 그림]]
{kind=link}
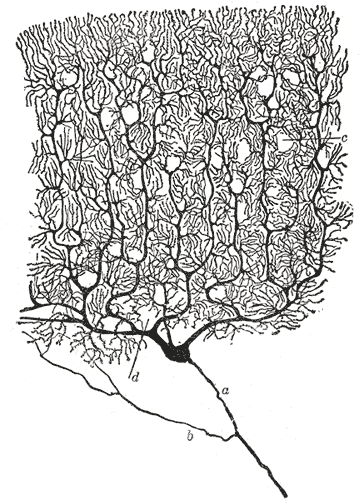
![골지 염색법이 미세한 세부 구조를 드러내는 능력을 보여주는, 산티아고 라몬 이 카할이 그린 [소뇌 피질 의 푸르킨예 세포 그림]]
{kind=link}
신경세포가 신경계의 기본 기능 단위라는 사실은 19세기 후반 스페인의 해부학자 산티아고 라몬 이 카할의 연구를 통해 처음으로 인정되었다.[^16]
개별 신경세포의 구조를 관찰할 수 있도록, 라몬 이 카할은 카밀로 골지가 개발한 은 염색 과정을 개선하였다.[^16] 개선된 과정은 "이중 침윤법"이라는 기술을 사용하며, 현재까지도 사용되고 있다.
1888년 라몬 이 카할은 조류 소뇌에 관한 논문을 발표하였다. 이 논문에서 그는 축삭과 수상돌기 사이의 문합(연결)에 대한 증거를 찾을 수 없었다고 밝히며, 각각의 신경 요소를 "자율적 단위(autonomous canton)"라고 불렀다.[^16][^17] 이것은 현대 신경과학의 핵심 교리 중 하나인 신경세포 교리(neuron doctrine)로 알려지게 되었다.[^16]
1891년, 독일의 해부학자 하인리히 빌헬름 발다이어는 신경세포 교리에 대한 매우 영향력 있는 리뷰를 작성하였으며, 여기서 신경계의 해부학적·생리학적 단위를 설명하기 위해 *뉴런(neuron)*이라는 용어를 도입하였다.[^71][^72]
은 침윤 염색은 신경해부학 연구에 유용한 방법인데, 아직 밝혀지지 않은 이유로 조직 내 소수의 세포만 염색하여, 다른 세포들과의 겹침 없이 개별 신경세포의 완전한 미세 구조를 드러내기 때문이다.[^18]
신경세포 교리
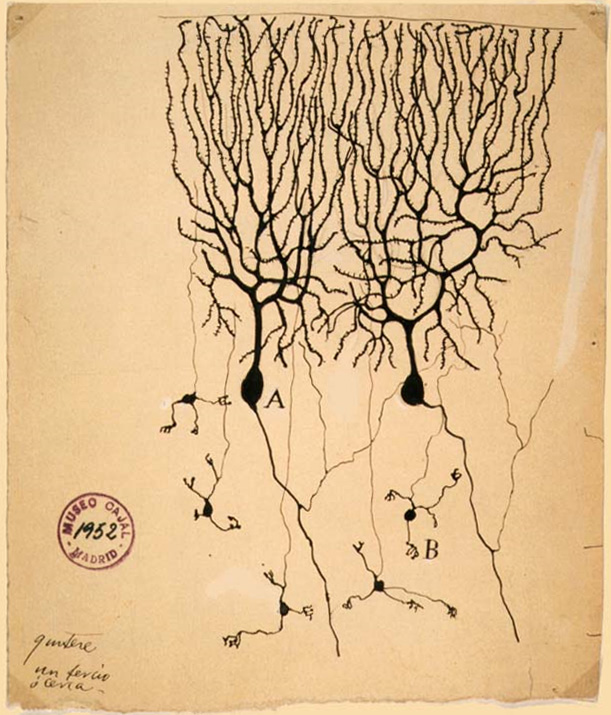
![1899년 스페인의 신경과학자 산티아고 라몬 이 카할이 그린 비둘기 [소뇌 의 신경세포 그림. (A)는 푸르킨예 세포를, (B)는 과립 세포를 나타내며, 둘 다 다극성이다.]] 신경세포 교리는 신경세포가 신경계의 기본적인 구조적·기능적 단위라는, 현재는 근본적인 개념이다. 이 이론은 19세기 후반 산티아고 라몬 이 카할에 의해 제안되었다. 이 이론은 신경세포가 (그물 구조로 연결된 것이 아닌) 분리된 세포이며, 대사적으로 독립된 단위로 작용한다고 주장하였다.
{kind=link}
이후의 발견들은 이 교리에 대한 수정을 가져왔다. 예를 들어, 비신경성인 신경교세포는 정보 처리에 필수적인 역할을 한다.[^73] 또한 전기적 시냅스는 이전에 생각했던 것보다 더 흔하며,[^74] 신경세포 간의 직접적인 세포질 연결을 구성한다. 사실, 신경세포는 더욱 긴밀한 결합을 형성할 수도 있는데, 오징어의 거대 축삭은 여러 축삭의 융합으로 형성된다.[^75]
라몬 이 카할은 또한 동적 극성의 법칙을 제안하였는데, 이는 신경세포가 수상돌기와 세포체에서 신호를 받아들이고, 활동전위의 형태로 축삭을 따라 세포체로부터 멀어지는 한 방향으로 전달한다는 것이다.[^19] 동적 극성의 법칙에는 중요한 예외가 있다. 수상돌기가 신경세포의 시냅스 출력 부위로 기능할 수 있으며,[^76] 축삭이 시냅스 입력을 받을 수도 있다.[^77]
신경세포의 구획 모델링
신경세포는 흔히 뇌의 "기본 단위"로 설명되지만, 내부적으로 연산을 수행한다. 신경세포는 수상돌기 내에서 입력을 통합하며, 신경세포를 기본 단위로 가정하는 모델에서는 이러한 복잡성이 상실된다. 수상돌기 가지는 공간적 구획으로 모델링할 수 있으며, 그 활동은 수동적 막 특성과 관련되지만, 시냅스로부터의 입력에 따라 달라질 수도 있다. 수상돌기의 구획 모델링은 전극으로 기록하기에는 너무 작은 신경세포의 행동을 이해하는 데 특히 유용한데, *초파리(Drosophila melanogaster)*의 경우가 이에 해당한다.[^78]
뇌의 신경세포
뇌의 신경세포 수는 종에 따라 극적으로 다르다. 인간의 경우, 대뇌피질에 약 100억~200억 개의 신경세포가 있는 것으로 추정되며 소뇌에는 550억~700억 개의 신경세포가 있다.[^20] 이에 비해, 선충류인 *예쁜꼬마선충(Caenorhabditis elegans)*은 단 302개의 신경세포만을 가지고 있어, 과학자들이 모든 신경세포를 매핑할 수 있었기 때문에 이상적인 모델 생물로 여겨진다. 생물학 실험에서 흔히 사용되는 초파리 *노랑초파리(Drosophila melanogaster)*는 약 100,000개의 신경세포를 가지고 있으며 많은 복잡한 행동을 보인다. 사용되는 신경전달물질의 종류에서 이온 채널 구성에 이르기까지, 신경세포의 많은 특성이 종 간에 유지되므로 과학자들은 더 복잡한 유기체에서 일어나는 과정을 훨씬 단순한 실험 체계에서 연구할 수 있다.
신경 질환
샤르코-마리-투스병(CMT)은 주로 발과 다리에서 시작하여 진행 단계에서 손과 팔로 확장되는 근육 조직 및 촉각 소실을 특징으로 하는 이질적인 유전성 신경 질환(신경병증)이다. 현재 치료법이 없는 이 질환은 가장 흔한 유전성 신경 질환 중 하나로, 100,000명당 36명에게 영향을 미친다.[^21]
알츠하이머병(AD)은 단순히 알츠하이머라고도 불리며, 일상생활 활동의 저하 및 신경정신과적 증상 또는 행동 변화와 함께 진행성 인지 저하를 특징으로 하는 신경퇴행성 질환이다.[^22] 가장 두드러진 초기 증상은 단기 기억 상실(기억상실증)로, 처음에는 가벼운 건망증으로 나타나다가 질병이 진행됨에 따라 점차 뚜렷해지며, 오래된 기억은 상대적으로 보존된다. 질환이 진행됨에 따라 인지(지적) 장애는 언어(실어증), 숙련된 동작(실행증), 인식(실인증) 영역으로 확장되며, 의사결정 및 계획과 같은 기능도 손상된다.[^23][^24]
파킨슨병(PD)은 단순히 파킨슨이라고도 불리며, 종종 운동 기능과 언어 능력을 저하시키는 중추신경계의 퇴행성 질환이다.[^25] 파킨슨병은 운동 장애라고 불리는 질환군에 속한다.[^79] 근육 강직, 떨림, 신체 움직임의 둔화(서동증), 그리고 극단적인 경우 신체 움직임의 상실(무동증)을 특징으로 한다. 주요 증상은 기저핵에 의한 운동 피질 자극의 감소로 인해 발생하며, 이는 일반적으로 뇌의 도파민 작동성 뉴런에서 생성되는 도파민의 불충분한 형성 및 작용에 의해 야기된다. 이차 증상으로는 고차원적 인지 기능 장애와 미묘한 언어 문제가 포함될 수 있다. 파킨슨병은 만성적이며 진행성이다.
중증 근무력증은 단순한 활동 시 근육의 변동성 약화와 피로감을 초래하는 신경근 질환이다. 근력 약화는 일반적으로 시냅스 후 신경근 접합부에서 아세틸콜린 수용체를 차단하는 순환 항체에 의해 발생하며, 이는 신경전달물질인 아세틸콜린의 자극 효과를 억제한다. 중증 근무력증은 면역억제제, 콜린에스테라아제 억제제, 그리고 선별된 경우에는 흉선절제술로 치료된다.
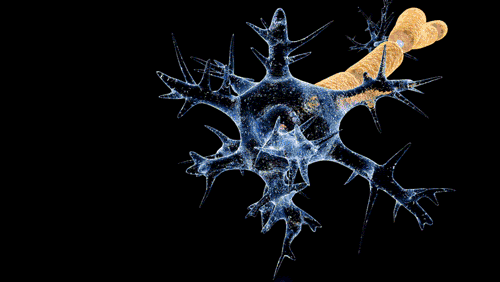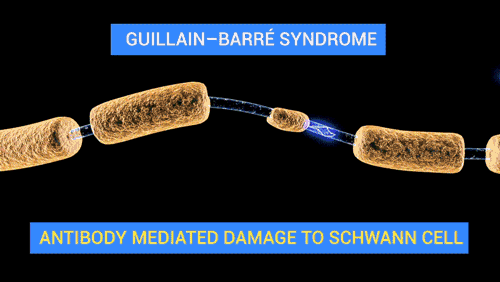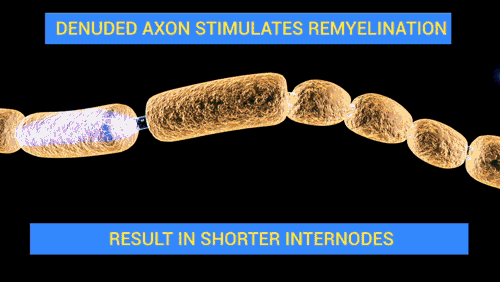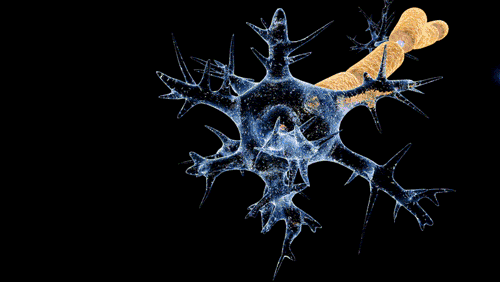
탈수초화
![[길랭-바레 증후군 – 탈수초화]] 탈수초화는 신경 섬유를 감싸고 있는 수초(미엘린)의 점진적인 소실을 특징으로 하는 과정이다. 수초가 손상되면 신경을 따른 신호 전도가 현저히 저하되거나 소실될 수 있으며, 신경은 결국 위축된다. 탈수초화는 중추신경계와 말초신경계 모두에 영향을 미칠 수 있으며, 다발성 경화증, 길랭-바레 증후군, 만성 염증성 탈수초성 다발신경병증 등 다양한 신경 질환에 기여한다. 탈수초화는 흔히 자가면역 반응에 의해 발생하지만, 바이러스 감염, 대사 장애, 외상 및 일부 약물에 의해서도 유발될 수 있다.
{kind=link}
축삭 변성
대부분의 손상 반응은 절단된 부분의 재봉합을 촉진하기 위한 칼슘 유입 신호를 포함하지만, 축삭 손상은 처음에 급성 축삭 변성을 초래하며, 이는 손상 후 30분 이내에 발생하는 근위부와 원위부 끝의 급속한 분리이다.[^26] 변성은 축삭막의 팽창과 함께 진행되며, 결국 구슬 모양의 형태를 형성한다. 축삭막 분해 이후 축삭 세포골격과 내부 세포소기관의 과립상 붕괴가 일어난다. 초기 변화로는 손상 부위의 결절 주변 영역에 미토콘드리아가 축적되는 것이 포함된다. 소포체가 분해되고 미토콘드리아가 부풀어 오르다가 결국 붕괴된다. 이러한 붕괴는 유비퀴틴과 칼파인 단백질분해효소(칼슘 이온의 유입에 의해 유발됨)에 의존하며, 이는 축삭 변성이 완전한 분절을 생성하는 능동적 과정임을 시사한다. 이 과정은 말초신경계에서 약 24시간이 소요되며, 중추신경계에서는 더 오래 걸린다. 축삭막 변성으로 이어지는 신호전달 경로는 아직 밝혀지지 않았다.
발달
신경세포는 신경줄기세포가 분열하여 분화된 신경세포를 생성하는 신경발생 과정을 통해 발달한다. 완전히 분화되면 더 이상 유사분열을 할 수 없다. 신경발생은 주로 배아 발달 과정에서 일어난다.
신경세포는 처음에 배아의 신경관에서 발달한다. 신경관은 뇌실대, 중간대, 변연대의 세 층으로 이루어져 있다. 뇌실대는 신경관의 중심관을 둘러싸며 뇌실막이 된다. 뇌실대의 분열하는 세포들은 중간대를 형성하며, 이 중간대는 연막층이라 불리는 신경관의 가장 바깥쪽 층까지 뻗어 있다. 뇌의 회백질은 중간대에서 유래한다. 중간대에 있는 신경세포의 돌기들이 변연대를 구성하며, 수초화되면 뇌의 백질이 된다.[^27]
신경세포의 분화는 크기에 따라 순서가 정해진다. 큰 운동신경세포가 먼저 분화한다. 더 작은 감각신경세포는 신경교세포와 함께 출생 시 분화한다.[^27]
성인 신경발생이 일어날 수 있으며, 인간 신경세포의 연령에 대한 연구에 따르면 이 과정은 소수의 세포에서만 일어나고 대뇌피질의 대다수 신경세포는 출생 전에 형성되어 교체 없이 유지된다. 성인 신경발생이 인간에게 어느 정도 존재하는지, 그리고 인지에 대한 기여도는 논쟁의 대상이며, 2018년에 서로 상충하는 보고서들이 발표되었다.[^80]
인체에는 신경세포로 분화할 수 있는 다양한 종류의 줄기세포가 존재한다. 연구자들은 "세포에 새로운 정체성을 강제로 부여하는" 직접교차분화를 통해 인간의 피부 세포를 신경세포로 변환하는 방법을 발견했다.[^28]
포유류 뇌의 신경발생 과정에서 전구세포와 줄기세포는 증식성 분열에서 분화성 분열로 진행된다. 이 진행 과정은 대뇌피질 층을 구성하는 신경세포와 신경교세포를 형성한다. 후성유전적 변형은 분화하는 신경줄기세포에서 유전자 발현을 조절하는 데 핵심적인 역할을 하며, 발달 중인 뇌와 성인 포유류 뇌에서 세포 운명 결정에 필수적이다. 후성유전적 변형에는 DNA 시토신 메틸화를 통한 5-메틸시토신 형성과 5-메틸시토신 탈메틸화가 포함된다.[^29] DNA 시토신 메틸화는 DNA 메틸전달효소(DNMT)에 의해 촉매된다. 메틸시토신 탈메틸화는 산화 반응(예: 5-메틸시토신에서 5-하이드록시메틸시토신으로)을 수행하는 TET 효소와 DNA 염기 절제 복구(BER) 경로의 효소에 의해 여러 단계에 걸쳐 촉매된다.[^30]
발달 중인 신경세포와 미세아교세포 간의 세포 간 소통 또한 적절한 신경발생과 뇌 발달에 필수적이다.[^81]
신경 재생
말초 축삭은 절단되어도 재성장할 수 있지만,[^31] 하나의 신경세포는 다른 유형의 신경세포로 기능적으로 대체될 수 없다(리나스의 법칙).[^7]
같이 보기
- 인공 신경세포
- 양방향 세포
- 생물학적 신경세포 모델
- 세포 신경과학
- 구획 신경세포 모델
- 커넥톰
- 도기엘 세포
- 성장원추
- 동물별 신경세포 수 목록
- 신경과학 데이터베이스 목록
- 신경세포 전류주성
- 신경가소성
- 적색 신경세포
- 숄 분석
더 읽을거리
-
-
-
-
-
-
외부 링크
-
-
-
-
-
- IBRO (국제 뇌 연구 기구). 특히 연구 자금이 부족한 국가들의 신경과학 연구를 지원.
- NeuronBank 신경 세포 유형과 시냅스 연결성을 목록화하는 온라인 신경체학 도구.
- 영장류 및 비영장류 뇌의 고해상도 신경해부학 이미지.
- 위키버시티의 신경과학부, 현재 두 개의 과정을 제공: 신경과학 기초 및 비교 신경과학.
- NIF 검색 – 뉴런 신경과학 정보 프레임워크를 통해 제공
- 세포 중심 데이터베이스 – 뉴런
- 뉴런 유형 전체 목록 NeuroLex에서 페틸라 협약에 따라 정리.
- NeuroMorpho.Org 신경 세포 형태의 디지털 재구성을 제공하는 온라인 데이터베이스.
- 면역조직화학 이미지 갤러리: 뉴런
- 칸 아카데미: 뉴런의 해부학
- 뉴런 이미지
참고 문헌
[^1]: Cite book title = Clinically Oriented Anatomy last1 = Moore first1 = Keith last2 = Dalley first2 = Arthur publisher = LWW year = 2005 isbn = 0-7817-3639-0 edition = 5th pages = [https://archive.org/de
[^2]: 중추신경계 재생을 위한 생체 내 성상세포-뉴런 재프로그래밍: 서술적 리뷰. (2023년 4월)
[^3]: 신경세관의 의학적 정의
[^4]: Al, Martini, Frederic Et. 해부학과 생리학' 2007 Ed.2007 Edition. Rex Bookstore, Inc.
[^5]: Betts, J Gordon. 해부학과 생리학. OpenStax CNX. (2023년 6월 8일)
[^6]: cite journal vauthors = Wilson NR, Runyan CA, Wang FL, Sur M title = 생체 내 개별 피질 억제 네트워크에 의한 나눗셈과 뺄셈 journal = Nature volume = 488 issue = 7411 pa
[^7]: cite journal vauthors = Llinás RR title = 포유류 뉴런과 중추신경계 기능의 고유 전기적 특성: 역사적 관점 journal = Frontiers in Cellular Neuroscience volume =
[^8]: cite journal vauthors = Ivannikov MV, Macleod GT title = 초파리 운동 신경 말단에서의 미토콘드리아 유리 Ca²⁺ 수준과 에너지 대사에 미치는 영향 journal = Biophysical Journal
[^9]: 왜 인간의 뇌는 이해하기 어려운가? 4명의 신경과학자에게 물었다.
[^10]: 미세아교세포는 특수화된 체세포 퓨린성 접합부를 통해 신경 기능을 모니터링하고 보호한다. (2020년 1월)
[^11]: Harris-Warrick, RM. 중추 패턴 생성기 네트워크에서의 신경조절과 유연성.. (2011년 10월)
[^12]: cite journal vauthors = Brown EN, Kass RE, Mitra PP title = 다중 신경 스파이크 트레인 데이터 분석: 최신 기술 및 미래 과제 journal = Nature Neuroscience volume = 7 issue =
[^13]: ''[[Oxford English Dictionary]]'', 제3판, 2003, ''s.v.''
[^14]: cite journal vauthors = Mehta AR, Mehta PR, Anderson SP, MacKinnon BL, Compston A title = 회백질 어원과 neuron(e) journal = Brain volume = 143 issue = 1 pages = 374–379 d
[^15]: Google Books Ngram Viewer
[^16]: cite journal vauthors = López-Muñoz F, Boya J, Alamo C title = 뉴런 이론, 신경과학의 초석, 산티아고 라몬 이 카할의 노벨상 수상 100주년을 맞아 journal = Brain
[^17]: Finger, Stanley. 신경과학의 기원: 뇌 기능 탐구의 역사. Oxford University Press
[^18]: cite journal vauthors = Grant G title = 1906년 노벨 생리의학상이 골지와 카할에게 공동 수여된 경위 journal = Brain Research Reviews volume = 55 issue = 2 pages =
[^19]: cite journal vauthors = Sabbatini RM date = 2003년 4월–7월 url = http://www.cerebromente.org.br/n17/history/neurons3_i.htm title = 뉴런과 시냅스: 발견의 역사 journal
[^20]: cite journal vauthors = von Bartheld CS, Bahney J, Herculano-Houzel S title = 인간 뇌의 실제 뉴런 및 신경아교세포 수 탐색: 150년간의 세포 계수 리뷰 jo
[^21]: cite journal vauthors = Krajewski KM, Lewis RA, Fuerst DR, Turansky C, Hinderer SR, Garbern J, Kamholz J, Shy ME title = 샤르코-마리-투스 질환에서의 신경학적 기능 장애와 축삭 변성
[^22]: 알츠하이머병에 대하여: 증상. 미국 국립노화연구소
[^23]: cite journal vauthors = Burns A, Iliffe S title = 알츠하이머병 journal = BMJ volume = 338 pages = b158 date = 2009년 2월 pmid = 19196745 doi = 10.1136/bmj.b158 s2cid = 8570
[^24]: cite journal vauthors = Querfurth HW, LaFerla FM title = 알츠하이머병 journal = The New England Journal of Medicine volume = 362 issue = 4 pages = 329–44 date = 2010년 1월 pmi
[^25]: 파킨슨병 정보 페이지. (2016년 6월 30일)
[^26]: cite journal vauthors = Kerschensteiner M, Schwab ME, Lichtman JW, Misgeld T s2cid = 25287010 title = 손상된 척수에서의 생체 내 축삭 변성 및 재생 영상화 journal
[^27]: Caire, Michael J.. 생리학, 시냅스. StatPearls Publishing. (2024)
[^28]: 인간 뉴런을 만드는 방법. (2011년 5월 26일)
[^29]: cite journal vauthors = Wang Z, Tang B, He Y, Jin P title = 신경 발생에서의 DNA 메틸화 역학 journal = Epigenomics volume = 8 issue = 3 pages = 401–14 date = 2016년 3월 pmid =
[^30]: 포유류 신경계 발달의 여러 단계에서 DNA 이중 가닥 절단 복구에 두 가지 DNA 복구 과정이 사용된다. 이러한 경로는 [[상동 재조합]]적 복구이며 u
[^31]: cite journal vauthors = Yiu G, He Z title = 중추신경계 축삭 재생의 신경아교세포 억제 journal = Nature Reviews. Neuroscience volume = 7 issue = 8 pages = 617–27 date = 2006년 8월 pmid
[^32]: Cambridge Dictionary, https://dictionary.cambridge.org/dictionary/english/neurone
[^33]: Najle, Sebastián R.. 초기 동물 진화에서 신경 유전자 발현 프로그램의 단계적 출현. (2023년 10월)
[^34]: Zayia LC, Tadi P. [https://www.ncbi.nlm.nih.gov/books/NBK554616/ 신경해부학, 운동 뉴런]. [2022년 7월 25일 업데이트]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023 Jan-.
[^35]: 신경계의 구성 요소는 무엇인가?. (2018년 10월)
[^36]: cite web first = Melissa last = Davies title = 뉴런: 크기 비교 url = https://www.ualberta.ca/~neuro/OnlineIntro/NeuronExample.htm work = 신경과학: 뇌를 통한 여행 dat
[^37]: cite web first = Eric H. last = Chudler name-list-style = vanc title = 뇌 사실과 수치 url = http://faculty.washington.edu/chudler/facts.html work = 어린이를 위한 신경과학 access-date =
[^38]: J.H. Smith, C. Rowland, B. Harland, S. Moslehi, K. Schobert, R.M. Montgomery, W.J. Watterson, J. Dalrymple-Alford, R.P. Taylor, "뉴런이 물리적 연결성을 극대화하기 위해 프랙탈 기하학을 활용하는 방법"
[^39]: cite journal vauthors = Herrup K, Yang Y title = 유사분열 후 뉴런의 세포 주기 조절: 모순어법인가 새로운 생물학인가? journal = Nature Reviews. Neuroscience volume = 8 issue = 5 pages
[^40]: Giménez, C.. [신경막의 조성과 구조: 생리학 및 병리학의 분자적 기초]. (1998년 2월)
[^41]: 주립 병원 회보. 정신이상위원회.
[^42]: cite journal vauthors = Zecca L, Gallorini M, Schünemann V, Trautwein AX, Gerlach M, Riederer P, Vezzoni P, Tampellini D title = 흑질에서의 철, 신경멜라닌 및 페리틴 함량
[^43]: cite journal vauthors = Herrero MT, Hirsch EC, Kastner A, Luquin MR, Javoy-Agid F, Gonzalo LM, Obeso JA, Agid Y title = 마카카 파시큘라리스의 카테콜아민성 뉴런에서 나이에 따른 신경멜라닌 축적
[^44]: cite journal vauthors = Brunk UT, Terman A title = 리포푸신: 연령 관련 축적 메커니즘과 세포 기능에 대한 영향 journal = Free Radical Biology & Medicine volume = 33 iss
[^45]: cite journal vauthors = Zhao B, Meka DP, Schellenberg R, König T, Schwanke B, Kobler O, Windhorst S, Kreutz MR, Mikhaylova M, Calderon de Anda F title = 미세소관이 신경 발생 동안 F-액틴 역학을 조절한다
[^46]: cite journal vauthors = Lee WC, Huang H, Feng G, Sanes JR, Brown EN, So PT, Nedivi E title = 성체 시각 피질의 GABA성 사이뉴런에서 수상돌기 수목의 동적 리모델링 journal = PLO
[^47]: cite journal vauthors = Gerber U title = 척추동물 망막의 대사성 글루타메이트 수용체 journal = Documenta Ophthalmologica. Advances in Ophthalmology volume = 106 issue = 1 pages =
[^48]: cite conference title = 쥐 망막 신경절 세포의 반복적 발화를 형성하는 이온 전도도 vauthors = Kolodin YO, Veselovskaia NN, Veselovsky NS, Fedulova SA conference = Ac
[^49]: 성체 쥐의 지속적으로 발화하는 망막 신경절 세포에서 흥분성의 기저가 되는 이온 전도도. Ykolodin.50webs.com. (2008-04-27)
[^50]: cite journal vauthors = Scammell TE, Jackson AC, Franks NP, Wisden W, Dauvilliers Y title = 히스타민: 신경 회로와 새로운 약물 journal = Sleep volume = 42 issue = 1 date = Janu
[^51]: Patch-seq 기술이 뇌에서 신경 세포의 변이를 묘사하는 데 도움을 준다. (2020년 12월 3일)
[^52]: Cite web last = Science AAAS title = 뇌 세포 인구조사 url = https://www.science.org/collections/brain-cell-census access-date = 2023-10-17
[^53]: Macpherson, Gordon. 블랙 의학 사전. Scarecrow Press. (2002)
[^54]: cite journal vauthors = Herculano-Houzel S title = 숫자로 보는 인간의 뇌: 선형적으로 확대된 영장류의 뇌 journal = Frontiers in Human Neuroscience volume = 3 page = 31 date = Nove
[^55]: cite journal vauthors = Drachman DA title = 우리에게 여분의 뇌가 있는가? journal = Neurology volume = 64 issue = 12 pages = 2004–5 date = 2005년 6월 pmid = 15985565 doi = 10.1212/01.WNL
[^56]: Ucar, Hasan. 수상돌기 가시 확장이 시냅스 전 엑소사이토시스에 미치는 기계적 작용. (2021년 12월)
[^57]: 연구자들이 뇌에서 새로운 유형의 세포 간 통신을 발견하다
[^58]: Schiapparelli, Lucio M.. 시각 시스템에서 연결된 뉴런 간의 다양한 단백질 수송에 대한 프로테오믹 스크리닝. (2022년 1월 25일)
[^59]: Levitan, Irwin B.. 뉴런. Oxford University Press
[^60]: O'Leary, Olivia F.. 미주 신경은 해마에서 BDNF 발현과 신경 발생을 조절한다. (2018년 2월 1일)
[^61]: cite web first = Eric H. last = Chudler name-list-style = vanc title = 신경과학 연구의 이정표 url = http://faculty.washington.edu/chudler/hist.html work = 어린이를 위한 신경과학 ac
[^62]: Patlak, Joe. 신경의 전기적 활동. (2000-11-01)
[^63]: 신경 시스템과 컴퓨터에서의 병렬 처리. North-Holland. (1990)
[^64]: cite book last1 = Eckert first1 = Roger last2 = Randall first2 = David name-list-style = vanc title = 동물 생리학: 메커니즘과 적응 year = 1983 publisher = W.H. Freeman
[^65]: Juusola, Mikko. 피질 네트워크에서의 스파이크 형태와 단계적 전위를 이용한 코딩. (2007년 2월)
[^66]: Zbili, Mickael. 수초화는 시냅스 전달의 아날로그 조절의 공간적 범위를 증가시킨다: 모델링 연구. (2020년 3월 3일)
[^67]: Zbili, M.. 시냅스 전달의 아날로그-디지털 조절의 과거와 미래
[^68]: 신경 코딩: 축삭에서의 아날로그 신호 전달
[^69]: Liu, Wenke. 나선 신경절 축삭에서의 활동 전위 미세 구조의 아날로그 전달
[^70]: Golgi, C. (1873). Sulla struttura della sostanza grigia del cervello (Comunicazione preventiva). ''Gaz. Med. Ital. Lomb.'' 33, 244–246. [https://books.google.com.br/books?id=cydbAAAAcAAJ].
[^71]: Finger, Stanley. 신경과학의 기원: 뇌 기능 탐구의 역사. Oxford University Press
[^72]: Whonamedit - 의학 명명법 사전
[^73]: cite journal vauthors = Witcher MR, Kirov SA, Harris KM title = 성숙 쥐 해마에서 시냅스 형성 동안 시냅스 주위 성상세포의 가소성 journal = Glia volume = 55 issue = 1
[^74]: cite journal vauthors = Connors BW, Long MA title = 포유류 뇌의 전기적 시냅스 journal = Annual Review of Neuroscience volume = 27 issue = 1 pages = 393–418 year = 2004
[^75]: cite journal vauthors = Guillery RW title = 시냅스 구조의 관찰: 뉴런 이론의 기원과 현재 상태 journal = Philosophical Transactions of the Royal Society o
[^76]: cite journal vauthors = Djurisic M, Antic S, Chen WR, Zecevic D title = 승모세포 수상돌기로부터의 전압 영상화: EPSP 감쇠와 스파이크 유발 영역 journal = The Journal of Neurosci
[^77]: cite journal vauthors = Cochilla AJ, Alford S title = 칠성장어 축삭에서의 글루타메이트 수용체 매개 시냅스 흥분 journal = The Journal of Physiology volume = 499 issue = Pt 2
[^78]: cite journal vauthors = Gouwens NW, Wilson RI title = 초파리 중추 뉴런에서의 신호 전파 journal = Journal of Neuroscience volume = 29 issue = 19 pages = 6239–6249 year = 2
[^79]: cite web title = 운동 장애 url = http://www.neuromodulation.com/movement-disorders work = 국제 신경조절 학회
[^80]: cite journal vauthors = Kempermann G, Gage FH, Aigner L, Song H, Curtis MA, Thuret S, Kuhn HG, Jessberger S, Frankland PW, Cameron HA, Gould E, Hen R, Abrous DN, Toni N, Schinder AF, Zhao X, Lucasse
[^81]: Cserép, Csaba. 체세포 퓨린성 접합부를 통한 신경 발달의 미세아교세포 조절. (2022년 9월)